 Lab Essentials
Lab Essentials AMPIVIEW® RNA probes
AMPIVIEW® RNA probes Enabling Your Projects
Enabling Your Projects  GMP Services
GMP Services Bulk Solutions
Bulk Solutions Research Travel Grant
Research Travel Grant Have You Published Using an Enzo Product?
Have You Published Using an Enzo Product?
- Easy-to-use kits (two-step) for screening SIRT5 inhibitors/activators; includes enough active enzyme for entire plate
- Includes optimal substrate selected from a panel of succinylated sites
- 96-well plate included, but can be adapted to higher well format
- Control inhibitors included
- Suitable for high-throughput screening (Z’-factors >0.73)
- Optimal, specific SIRT5 substrate means low enzyme concentration, making ‘hit’ validation easy
- While the FLUOR DE LYS®–Succinyl substrate (Prod. No. BML-KI590, in FLUOR DE LYS® SIRT5 Fluorometric Drug Discovery Assay Kit, Prod. No. BML-AK413) contains an AMC (‘blue’) fluorophore that uses commonly-used wavelengths, FLUOR DE LYS®–Succinyl Green substrate has longer excitation/emission wavelengths, avoiding interference often seen with screening compounds at shorter wavelengths.
The FLUOR DE LYS® Green SIRT5 Fluorometric Drug Discovery Kit is a complete assay system designed to measure the lysyl desuccinylase activity of the recombinant human SIRT5 included in the kit. A black 96-well microplate is packaged with the kit, but it should be noted that reagents of the FLUOR DE LYS® system have also been successfully employed in other formats, including cuvettes and 384-well plates.
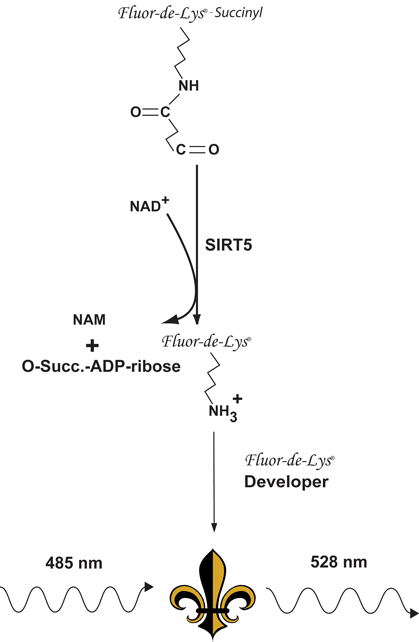
The Green SIRT5 Fluorescent Activity Assay is based on the unique FLUOR DE LYS®-Succinyl GreenSubstrate/Developer combination. The assay procedure has two steps. The FLUOR DE LYS®-Succinyl GreenSubstrate, which comprises a lysine residue, Nε-succinylated on its side-chains, is first incubated with human recombinant SIRT5 together with the cosubstrate NAD+. Desuccinylation of FLUOR DE LYS®-Succinyl Green sensitizes it so that, in the second step, treatment with the FLUOR DE LYS® Developer produces a fluorophore. Use of a succinylated, rather than acetylated substrate with SIRT5 results in readily observed saturation kinetics and a greater than 1000-fold increase in assay sensitivity
Shipping: Available products typically ship within 24/48h, via priority shipping.
Do you need support? Contact Customer Service or Technical Support.
Online Account
Access or Create Your Account
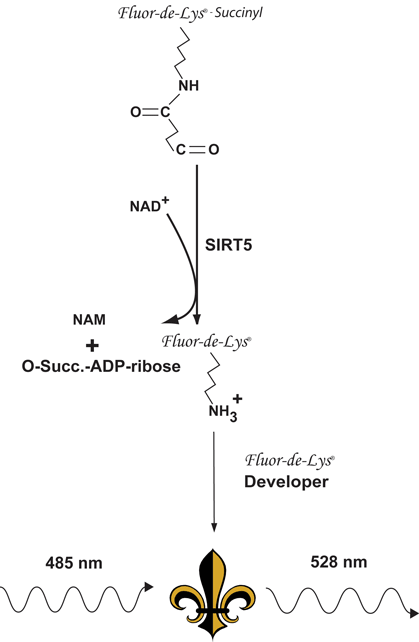
Reaction Scheme of the SIRT5 Fluorescent Activity Assay*. NAD+-dependent desuccinylation of the substrate by recombinant human SIRT5 sensitizes it to Developer, which then generates a fluorophore (symbol). The fluorophore is excited with 485 nm light and the emitted light (528 nm) is detected on a fluorometric plate reader. NAD+ is consumed in the reaction to produce nicotinamide (NAM) and O-succinyl-ADP-ribose.
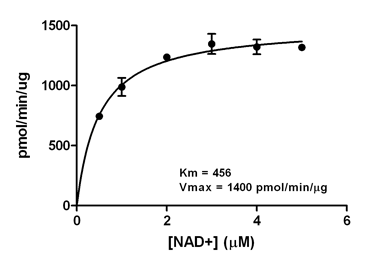
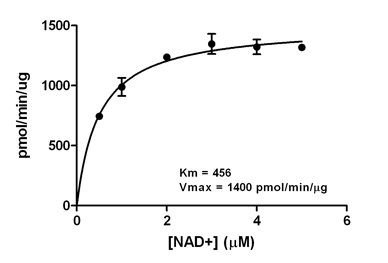
Dependence of SIRT5 Kinetics on the Concentration of NAD+. Initial desuccinylation rates of SIRT5 (10 ng) were determined with 20 min. incubations (37°C) in the presence of 0.5 mM FLUOR DE LYS®-Succinyl Green and the indicated concentrations of NAD+. Reactions were stopped with FLUOR DE LYS® Developer/2 mM nicotinamide and fluorescence measured. Each point represents the mean of three determinations and the error bars are standard deviations. The line is a non-linear least-squares fit to the Michaelis-Menten equation. The Km for NAD+ was 456 µM and the Vmax was 1400 pmol/min/µg.
| Regulatory Status |
RUO – Research Use Only |
|---|
Related Products
FLUOR DE LYS® HDAC fluorometric activity assay kit
BML-AK500
First-to-market by the leader in Epigenetics research tools

| Application | Activity assay, Fluorescent detection, HTS |
|---|
COLOR DE LYS® HDAC colorimetric activity assay kit
BML-AK501
Simplify HDAC activity analysis with the fastest colorimetric assay on the market
| Application | Activity assay, Colorimetric detection, HTS |
|---|
| Application | Activity assay, Cell-based assays, Fluorescent detection, HTS |
|---|
| Application | Activity assay, Cell-based assays, Fluorescent detection, HTS |
|---|
Last modified: May 29, 2024