 Lab Essentials
Lab Essentials AMPIVIEW® RNA probes
AMPIVIEW® RNA probes Enabling Your Projects
Enabling Your Projects  GMP Services
GMP Services Bulk Solutions
Bulk Solutions Research Travel Grant
Research Travel Grant Have You Published Using an Enzo Product?
Have You Published Using an Enzo Product?
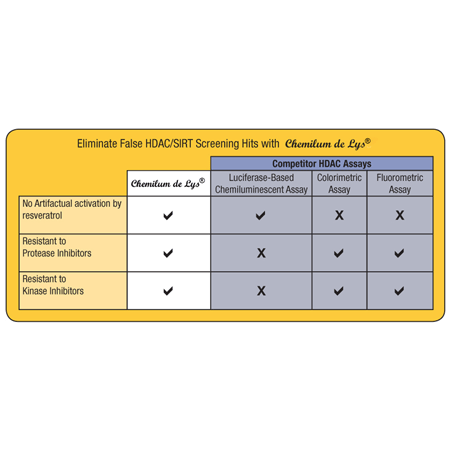
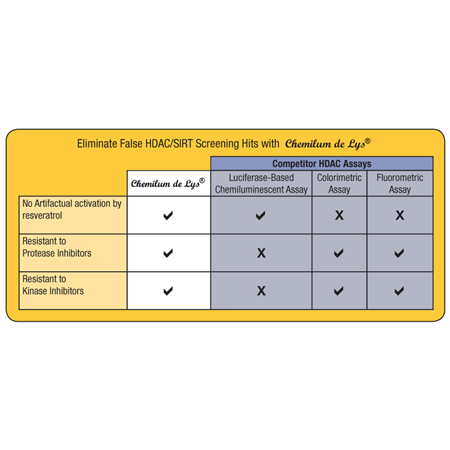
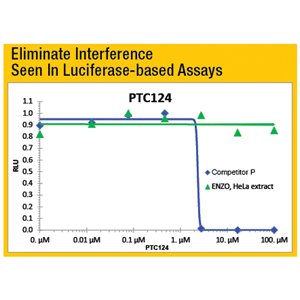
- High Specificity assay eliminates false positives or negatives
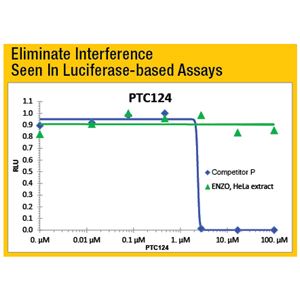
- Superior Signal-to-Noise ratio with no interference from cell extract detergents
- Delivers consistent results from a validated system
The HDAC/SIRT Chemiluminescent Drug Discovery Kit is a complete assay system designed to measure histone deacetylase (HDAC) and sirtuin activity in cell or nuclear extracts, immunoprecipitates or purified enzymes. It comes in a convenient 96-well format, with all reagents necessary for chemilumnescent HDAC activity measurements and calibration of the assay. In addition, a HeLa nuclear extract, rich in HDAC activity, is included with the kit. The extract is useful as either a positive control or as the source of HDAC activity for inhibitor/drug screening. Also included are Trichostatin A and Nicotinamide, which may be used as model inhibitors for HDACs and sirtuins, respectively.
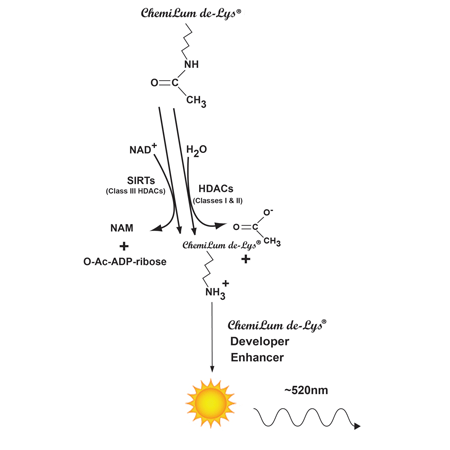
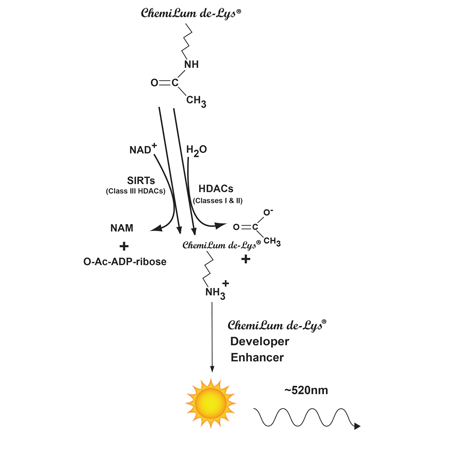
The HDAC Chemiluminescent Activity Assay is based on the unique CHEMILUM DE LYS® Substrate and Developer combination.
The CHEMILUM DE LYS® system (Chemiluminescent Histone deAcetylase Lysyl Substrate/Developer) is a highly sensitive and convenient alternative to radiolabeled, acetylated histones or peptide/HPLC methods for the assay of histone deacetylases. The assay procedure has three steps (Fig. 1). First, the CHEMILUM DE LYS® Substrate, which comprises an acetylated lysine side chain, is incubated with a sample containing HDAC activity (HeLa nuclear or other extract, purified enzyme, bead-bound immunocomplex, etc.). Deacetylation of the substrate sensitizes the substrate so that, in the second step, treatment with the CHEMILUM DE LYS® Developer followed by Enhancer produces light. The reaction is luciferase-free
Shipping: Available products typically ship within 24/48h, via priority shipping.
Do you need support? Contact Customer Service or Technical Support.
Online Account
Access or Create Your Account
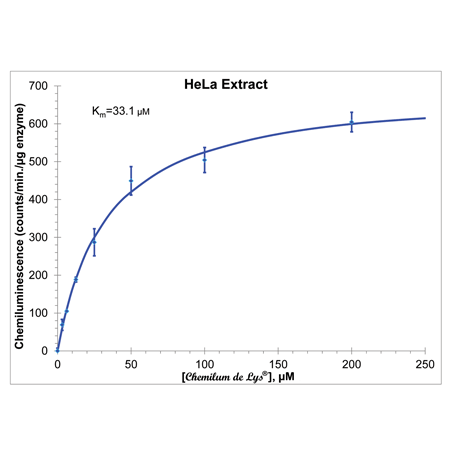
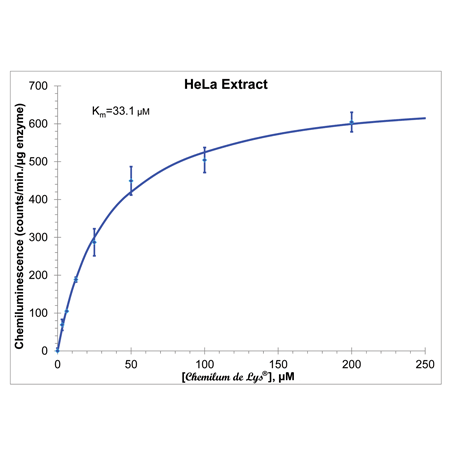
Kinetics of CHEMILUM DE LYS® Substrate Deacetylation by HeLa HDAC Activity. HeLa Nuclear extract (0.35 µl/7 µg per well) was incubated (37°C) with indicated concentrations of substrate. Reactions were stopped after 10 min. with CHEMILUM DE LYS® Developer, incubated for 120 minutes, then Enhancer was added and chemiluminescence measured (BMG Labtech, FluoStar Optima). Points are the mean of three determinations and error bars are standard errors of the means. Each determination is the average of at least five chemiluminescence readings taken in the first ten minutes. Kinetic parameters and the line derive from a non-linear least squares fit of the data to the Michaelis-Menten equation (Microsoft XL, Solver tool).
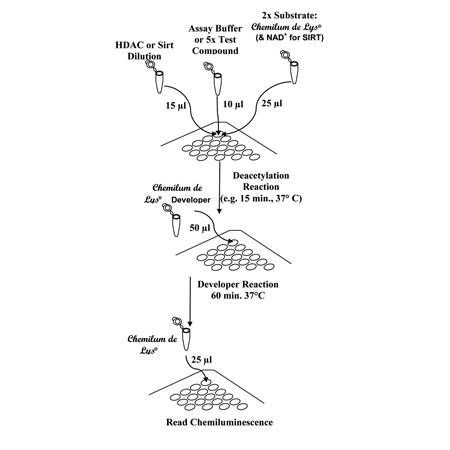
Performing the CHEMILUM DE LYS® HDAC Activity Assay. The procedure is done in three stages. First, the components of the deacetylation reaction (HeLa extract, buffer or test compound, substrate) are mixed. Following an incubation in which substrate deacetylation takes place, Developer is added and mixed. This stops the deacetylation and produces a product that will generate a chemiluminescent signal when enhancer is added. The signal can be read in 60 min, after the addition of Enhancer. The scheme depicts mixes for “Control” or “Test Sample” reactions. When performing the assay on 384 well plates, all volumes should be cut in half.
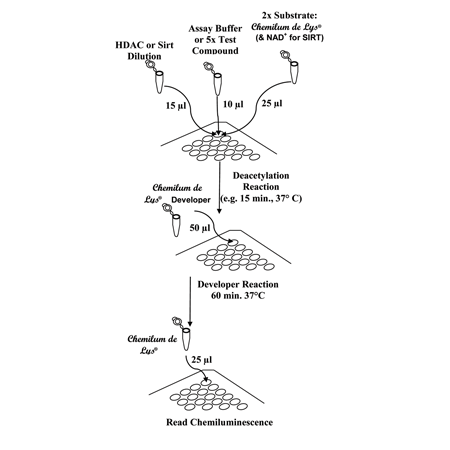
Performing the Chemilum de Lys HDAC Activity Assay. The procedure is done in three stages. First, the components of the deacetylation reaction (HeLa extract, buffer or test compound, substrate) are mixed. Following an incubation in which substrate deacetylation takes place, Developer is added and mixed. This stops the deacetylation and produces a product that will generate a chemiluminescent signal when enhancer is added. The signal can be read in 60 min, after the addition of Enhancer. The scheme depicts mixes for “Control” or “Test Sample” reactions. When performing the assay on 384 well plates, all volumes should be cut in half.
CHEMILUM DE LYS® HDAC/SIRT Chemiluminescent drug discovery kit Fig8
CHEMILUM DE LYS® HDAC/SIRT Chemiluminescent drug discovery kit Fig1
| Regulatory Status |
RUO – Research Use Only |
|---|
Related Products
FLUOR DE LYS® HDAC fluorometric activity assay kit
BML-AK500
First-to-market by the leader in Epigenetics research tools

| Application | Activity assay, Fluorescent detection, HTS |
|---|
COLOR DE LYS® HDAC colorimetric activity assay kit
BML-AK501
Simplify HDAC activity analysis with the fastest colorimetric assay on the market
| Application | Activity assay, Colorimetric detection, HTS |
|---|
FLUOR DE LYS® HDAC fluorometric cellular activity assay kit
BML-AK503
Monitor HDAC activity in cell culture using cell-permeable FLUOR DE LYS® substrate
| Application | Activity assay, Cell-based assays, Fluorescent detection, HTS |
|---|
| Application | Activity assay, Cell-based assays, Fluorescent detection, HTS |
|---|
Last modified: May 29, 2024